
Advances in Animal and Veterinary Sciences


Research Article
Advances in Animal and Veterinary Sciences 1 (5): 143 – 147DNA Prime and Protein Boost Immunization with Combined SOD–L7/L12 Antigen Confers Protection to Mice against Brucella Abortus 544 Challenge
Pallab Chaudhuri*, Harisankar Singha, Tapas Kumar Goswami, Candrakanta Jana, Gunjan Shukla
-
Division of Bacteriology and Mycology Indian Veterinary Research Institute, Izatnagar –243122
*Corresponding author:[email protected]
ARTICLE CITATION:
Chaudhuri P, Singha H, Goswami TK, Jana C and Shukla G (2013). DNA prime and protein boost immunization with combined SOD–L7/L12 antigen confers protection to mice against Brucella abortus 544 challenge. Adv. Anim. Vet. Sci. 1 (5): 143 – 147.
Received: 2013–09–02, Revised: 2013–09–26, Accepted: 2013–09–26
The electronic version of this article is the complete one and can be found online at
(
http://nexusacademicpublishers.com/table_contents_detail/4/103/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Brucella abortus is an intracellular pathogen that has zoonotic implication. Currently used live attenuated vaccines have several limitations including interference in serodiagnosis of clinical infection and presence of residual virulence in the vaccine strains. There is an urgent need to develop a safe vaccine for control of brucellosis. Here, we describe development of a DNA vaccine construct by combining two immunodominant antigens namely, copper–zinc superoxide dismutase (Cu–Zn SOD) and L7/L12 ribosomal protein. Both the genes were amplified and ligated together to produce the DNA vaccine construct. The combine gene was further expressed in prokaryotic system to produce recombinant fusion protein. A DNA prime–protein boost strategy was followed to immunize mice. Strong immune response dominated by IgG2a subtype shows immune skewing towards Th1 type. Further we have showed that combined antigen conferred significantly higher level of protection against a challenge infection as compare to single antigen immunization. However, the level of protection conferred by combined antigen was lower that the S19 live vaccine. Our report shows the potential of SOD: L7/L12 combined antigen as candidate vaccine eliciting strong immune response against experimental murine brucellosis.
INTRODUCTION
Brucellae are gram–negative, facultative intracellular microorganism. Species in the genus Brucella are the causative agents of brucellosis and the disease is endemic in many countries. It causes abortion in pregnant animals, resulting from complex, not well understood interactions between the tissues of placenta, the intracellular Brucellae and the fetus (Ko and Splitter, 2003). Orchitis and vesiculitis are the major signs and common manifestations in infected bulls (Corbel, 1997). Because of its intracellular localization, cell mediated immune response play important role in resolving infection. Brucella has been categorized as a potential agent for bio–terrorism because it is highly infectious to human and there is no effective human vaccine for brucellosis.
Currently, B.abortus strain 19 and RB51 are being used for cattle and Rev1 for small ruminants to control animal brucellosis in various countries (Cardena et al., 2009; Jimenez de Bagues et al., 1994; Schurig et al., 1991; Wyckoff et al., 2005). In India, calf hood vaccination is practiced using B. abortus stain 19 for control of brucellosis in cattle and buffaloes. However, this vaccine suffers from some serious drawbacks like – residual virulence for animals and human, interference in sero–diagnosis of clinical infection (Nicoletti, 1990). The identification of Brucella antigens with the capacity to elicit protective immune response is of great interest for the development of sub–cellular vaccines that do not have the drawbacks of live attenuated vaccine. Several Brucella antigens have been evaluated for their immunogenic properties (Al–Mariri A, Tibor et al., 2001; Bachrach et al., 1994; Cassataro et al., 2005; Mallick et al., 2007; Oliveira and Splitter, 1996; Oliveira et al., 1992; Oliveira et al., 1994; Oliveira et al., 1996; Oliveira and Splitter, 1994; Tabatabai and Pugh,1994; Vemulapalli et al., 2000). Of these, Brucella abortus L7/L12 ribosomal protein has been identified as an immunodominant antigen (Brooks–Worrel and Splitter, 1992). This protein is a major constituent of brucellin which elicits delayed –type hypersensitivity (Bachrach et al., 1994).The other important Brucella antigen is Cu–Zn SOD. Brucella Cu–Zn SOD is a 18.5–19 kDa protein encoded by 522 bp open reading frame of sodC gene (Bricker et al., 1990). Cu–Zn SOD is one of the principal antioxidants of B. abortus and plays a vital role in defense versus the respiratory burst of the macrophage (Tatum et al., 1992). Several reports have suggested that DNA, RNA or synthetic peptide based Brucella Cu–Zn SOD vaccine can proficiently induce a Th1 type of immune response and some level of protection against challenge infection in mice (Tabatabai and Pugh,1994; Onate et al., 2003; Munoz–Montesino et al., 2004; Onate et al., 2005).
In this report we describe the development of a chimeric DNA vaccine construct containing Cu–Zn SOD and L7/L12 ribosomal gene and expression of both the genes as SOD:L7/L12 fusion protein. We further evaluate the protective efficacy of DNA vaccine and the fusion protein against virulent challenge in experimental animal, mice.
MATERIALS AND METHODS
Animals
Inbred female Swiss albino mice (4–6 weeks old), weighing approximately 20 g, were obtained from the Laboratory Animal Resources, Indian Veterinary Research Institute, Izatnagar, India. Animal experiments were carried out after obtaining approval from the Institutional animal ethics committee. All the mice were housed under standard conditions at the animal shed facilities and provided sufficient water and food.
Bacterial Strain and Vector
The virulent Brucella abortus Strain 544 and the attenuated Brucella abortus Strain–19 was obtained from Division of Standardization, Indian Veterinary Research Institute (IVRI), Izatnagar, India. Escherichia coli DH5µ strain (Life technology, USA) was used as host for cloning of DNA vaccine construct whereas E. coli M15 strain (Qiagen, USA) was used for expression of SOD:L7/l12 fusion protein. Prokaryotic expression vector pQE–30 (Qiagen, USA) and eukaryotic expression vector pVAX1 (Invivogen, CA, USA) were used in the present study. Brucella cells were grown on Brucella broth under aerobic condition for 72 hours at 37oC and E. coli cells were grown in Luria –Bartani (LB) medium, supplemented with required antibiotics (kanamycin and ampicillin), under aerobic condition at 37oC.
Construction of pVSL DNA Vaccine:
Vector pVAX1 was used to develop chimeric DNA vaccine construct encompassing Cu–Zn SOD (sodC gene) and L7/L12 ribosomal protein (l7/l12 gene) of Brucella abortus. A 522 bp DNA fragment encoding sodC gene and 375 bp l7/l12 genes were amplified from Brucella abortus strain 544 DNA by Polymerase Chain Reaction. The primer sequences of the sodC were 5'–CGGCGGATCCACCATGGGAAAGTCCTTATTTATTGCATC–3' (primer 1 with BamH I site underlined) and 5'–CGGCGTCGACTTCGATCACGCCGCAGGCAAAACG–3' (primer 2 with Sal I site underlined). The primer sequences of the l7/l12 gene were 5'–CGCCGTCGACTCGTCCGGCGGAGGCGGA–3' (primer 3 with sal I site underlined) and 5'–GCCCCTGCAGTTACTTGAGTTCAACCTTGGC–3' (primer 4 with Pst I site underlined). The PCR amplified sodC gene, without stop codon, was placed to upstream of l7/l12 gene product and ligated to pVax1 vector. Two genes were separated by a nucleotide spacer that encoded for six amino acids (2 serine and 4 glycin). The resulting plasmid was designated as pVSL. The DNA vaccine construct, pVSL, was transformed into chemically (CaCl2) competent E. coli DH5 α cells by heat shock method. Recombinant E. coli harbouring DNA vaccine construct was cultured in LB medium supplemented with kanamycin (25 μg/mL) for large scale production of DNA vaccine, pVSL. DNA vaccine constructs for individual genes, l7/l12 and sodC, were prepared in pVax1 vector and the DNA vaccine constructs were named as pVL7 /L12 and pVsod, respectively.
Expression and Purification of Sod: L7/L12 Fusion Protein
To generate recombinant SOD:L7/L12 fusion protein, the target fusion gene was cleaved from the pVSL plasmid by restriction endonucleases BamH I and Pst I. The released insert was ligated to pQE–30 expression vector and transformed into chemically competent E.coli M15 cells. Recombinant E. coli cells were grown in LB broth supplemented with kanamycin (25 μg/mL) and ampicillin (100 μg/mL) and IPTG (1mM) was used for induction of expression. Purification of SOD:L7/L12 fusion protein was made using Ni2+–NTA agarose resin (Qiagen, USA). Purified protein was identified by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE). Recombinant protein for individual genes, l7/l12 and sodC was prepared using prokaryotic expression vector.
Sequencing of pVSL
The cloned L7/L12 ribosomal protein gene and sodC gene in pVSL was sequenced by dideoxy chain termination method. The nucleotide sequence was analyzed by using ‘EditSeq’ program of Lasergene software (DNASTAR, Madison, USA).
Immunization
The immunization studies were carried out in female Swiss albino mice. Each group was consisted of 15 animals. The mice were inoculated intramuscularly with 100 µg of pVSL, pVL7/L12 or pVsod in 50 µL of PBS. A booster immunization was carried out on day 21 with 50 µg of recombinant fusion protein (SOD:L7/L12), L7/L12 or Cu–Zn SOD. A group of mice was immunized with PBS as negative control. As positive control, group of mice was inoculated intraperitoneally with 1 x 105 CFU of B. abortus strain 19 in 0.2 mL of PBS. Five mice from each experimental group were bled for collection of sera samples at different time intervals (0, 20, 35 and 50th day).
Analysis of Specific Antibody Responses by Elisa
To assess the antibody response to SOD:L7/L12 fusion protein, sera samples were collected at different time points from the immunized mice. Total IgG and IgG isotype response were determined by indirect ELISA. Immunoplates (Maxisorp, Nunc) were coated with purified recombinant SOD:L7/L12 protein at a concentration of 2.5 µg/mL (100 µL/well ) diluted in carbonate–bicarbonate buffer (0.05M, pH 9.6). Goat anti–mouse IgG–HRPO conjugate (1:2500) was used as secondary antibody. ELISA plates were developed in 100 µL of freshly prepared substrate solution (200 µmol o–phenylene–diamine and 0.04% H2O2 in citrate buffer) and the reaction was stopped by addition of 50 µL of 0.18 M sulfuric acid per well. Absorbance of the developed color was measured at 492 nm. The titer of each serum was expressed as A492 value and normalized to value of control sera tested at 1:50 dilution. Similarly, titer of IgG isotypes were determined by using goat anti–mouse IgG1, IgG2a and IgG2b– HRPO conjugates (Santa Cruz, USA) at a dilution of 1:10,000. Mean values and standard deviations were calculated to depict immunoglobulin response to individual vaccine group.
Protection Study against Brucella abortus 544 Challenge
Four weeks after last vaccination, mice were challenged with a virulent culture of Brucella abortus 544 (1 X 105 CFU / mice) by intraperitoneal route in 0.2 mL PBS. After 30 days of challenge, mice from each group were euthanized; spleens were aseptically removed and triturated with 2 mL of 1.5M PBS (pH7.4) with 0.1% TritonX–100 using sterile sand. Ten–fold serial dilutions of spleen cell lysate were made and 50 µL from each dilution was plated on Typtose Phosphate Broth (TPB) medium with 1.5% agar in triplicate. For S–19 vaccinated mice group, 0.1% Erythriotol was added in the plate for differentiating S–19 and Brucella abortus 544. Plates were further incubated at 37oC with 10% CO2 pressure. Colonies appeared after three days of incubation were counted and calculated as log10 colony forming unit (cfu) per spleen. Protection conferred by individual vaccine group was expressed as log10 unit of protection and it was derived by subtracting mean log10 cfu for the experimental group from the negative control group.
RESULTS
Construction of DNA Vaccine and Expression of Fusion Protein
The construction of pVSL DNA vaccine encoding Cu–Zn SOD and L7/L12 ribosomal protein genes was confirmed by PCR (Figure 1A). Analysis of DNA sequence showed correct orientation and also sequence similarity of both the genes (data not shown). The gene insert was further released from pVSL and subcloned into pQE–30 prokaryotic expression vector. Expressed SOD: L7/L12 fusion protein showed the desired protein band on SDS–PAGE (Figure 1. To ensure antigen expression from pVSL DNA vaccine construct in a eukaryotic system, Vero cells were transfected (TransFast transfection ragent, Promega, USA) and further immunofluorescence was detected by using SOD:L7/L12 specific antiserum (data not shown).
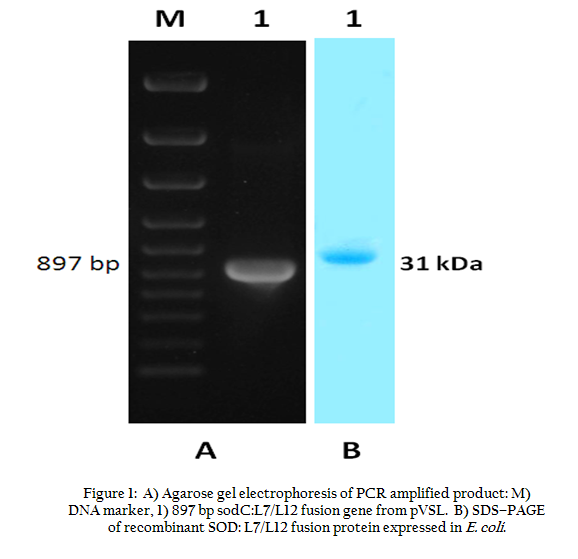
Figure 1: A) Agarose gel electrophoresis of PCR amplified product : M) DNA marker, 1) 897 bp sodC:L7/L12 fusion gene from pVSL. B) SDS–PAGE of recombinant SOD:L7/L12 fusion protein expressed in E. coli.
Analysis of Immune Response in Mice Immunized with DNA Vaccines
In the present study we followed DNA prime–protein boost regimen for immunization of mice. Sera collected from 5 mice of individual vaccine group were analyzed by indirect ELISA (Figure 2). The detectable antibody titer was measurable on 35th day sera of pVSL group. The level of antibody titer was maintained even on day 50 post immunization (Figure 2).
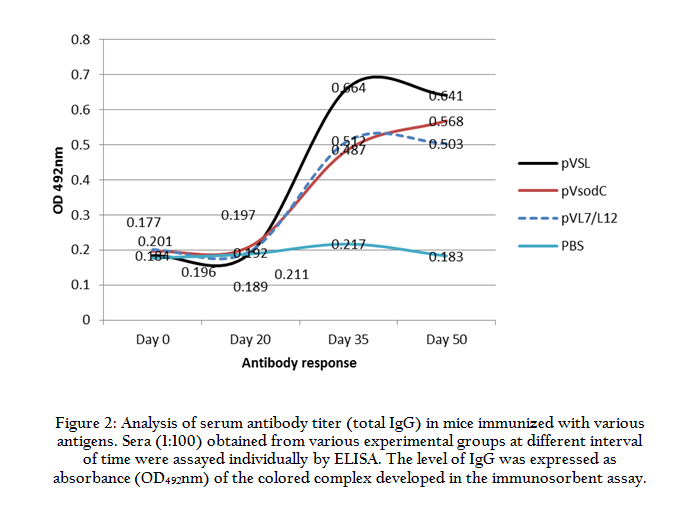
Figure 2: Analysis of serum antibody titer (total IgG) in mice immunized with various antigens. Sera (1:100) obtained from various experimental groups at different interval of time were assayed individually by ELISA. The level of IgG was expressed as absorbance (OD492nm) of the colored complex developed in the immunosorbent assay.
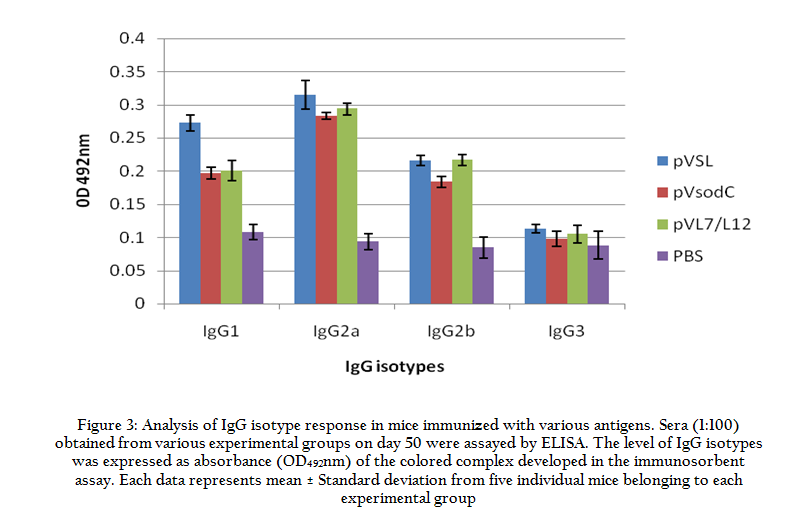
Figure 3: Analysis of IgG isotype response in mice immunized with various antigens. Sera (1:100) obtained from various experimental groups on day 50 were assayed by ELISA
Protective Efficacy of DNA Vaccine in Mice
Degree of protection conferred by individual vaccine group was assessed by calculating the log10 value of bacterial load in the spleen. Immunization with pVSL resulted in a significantly higher degree of protection in comparison to pVL7/L12 or pVSOD alone (Table 1). However, the level of protection conferred by pVSL was lesser than S19 live vaccine.
Table 1: Level of protection against Brucella abortus 544 infection conferred by various vaccine antigens in mice on day 30 post challenge
DISCUSSIONS
Recent trend in the development of successful vaccine candidate against various intracellular pathogens emphasizes on importance of simultaneous activation of cellular immunity along with desirable humoral counterpart. The protective immunity against brucellosis, a continuous threat to our livestock population, relies more on Th1 lymphocytes that play a prominent role in stimulation of cell–mediated immune response. The subunit vaccines have advantages as they exclude safety concerns associated with live vaccines, These subunit vaccines in the form of DNA vaccine or purified proteins elicit immune responses in animals. In order to introduce an improved vaccine against Brucella infection, subunit vaccines have been used but not found to be efficient enough in protecting animals from subsequent infection and disease (Singha et al., 2008; Winter and Rowe, 1988; Confer, 1987).
A number of proteins have been studied in laboratory animal model which could impart protective immunity to brucellosis (Al–Mariri A, Tibor et al., 2001; Tabatabai and Pugh,1994; Bachrach et al., 1994; Oliveira and Splitter, 1996; Vemulapalli et al., 2000). L7/L12 ribosomal protein and Cu–Zn SOD are among the immuno–dominant antigens that have shown to trigger immune system of the host with modest response (Bachrach et al., 1994; Mallick et al., 2007; Munoz–Montesino et al., 2004; Oliveira and Splitter, 1994; Singha et al., 2008; Brooks–Worrel and Splitter, 1992). To improve the efficacy of DNA vaccines, prime–boost regimen has been followed in many studies (Cassataro et al., 2007; Zhang et al., 2012). This is based on the use of DNA to prime the host immune system followed by boosting the immune system with protein antigen. Our present study is mainly focused on the possible use of two immune–dominant antigens as fusion product to be delivered as DNA vaccine as well as fusion protein. There was no detectable antibody specific to either L7/L22 or Cu–Zn SOD after primary immunization with DNA vaccine constructs. However, after booster immunization with respective protein antigen, there was considerable production of antigen specific antibodies. DNA vaccines are found poor immunogen in eliciting antigen specific humoral response. We observed that the prime–boost immunization regimen was effective to elicit both cellular and humoral immune response to Brucella antigens.
We have demonstrated that combined antigen when delivered as DNA vaccine followed by protein booster conferred significantly higher protective immunity to mice against virulent challenge in comparison to individual antigen. We have shown that the linkage between sodC and l7/l12 genes have synergistic effect, mounting strong immune response. In the present study we have observed that primary immunization with DNA vaccine constructs did not elicit detectable antibody production in immunized mice. Antigen specific antibodies were produced only after booster immunization with protein antigens. Induced antibody response was characterized by high titers of SOD:L7/L12 specific IgG in pVSL vaccinated mice. Injection with individual antigen also increased the production of IgG antibodies. However, the increased level was significantly lower than combined vaccine. Similarly, S–19 vaccine group has also showed high level of IgG induction on that in turn ensured to maximum protective efficacy against infection. During generation of bivalent DNA vaccine construct, we have included a spacer sequence of six amino acids (two serine and four glycin) placed between the two genes, sodC and l7/l12. Presence of spacer sequence would facilitate both the protein antigens of fusion protein to show protein folding as independent domain.
To establish vaccine potential of chimeric DNA vaccine construct, bacterial burden in spleen of the immunized mice were enumerated. After 30 days of virulent challenge with B. abortus 544 strain, mice immunized with combined antigen showed considerably higher level of protection in comparison to single antigen immunized mice group. We also observed splenomegaly in certain animals challenged with Brucella abortus 544 infection . Animals exposed to infection without prior immunization with either DNA vaccine or S–19 vaccine showed prominent splenomegaly. Moreover, discoloration of surface, granuloma formation and superficial multiple foci, were also noted in the spleen, collected from control group of mice. In contrast, there were no macroscopic lesions observed in the DNA or S19 vaccinated mice.
Immunized animals with higher IgG2a titer showed the higher level of protection on individual level. It is important that Fc portion of IgG2a binds to the Fc receptors present on the surface of phagocytes which in turn activates a wide spectrum of antimicrobial response like release of various inflammatory mediators, cytokine synthesis, generation of reactive oxidant species (Unkeless et al., 1998). There was a good correlation between increased level of IgG2a and containment of the infection in the immunized animals. We do have observed that pVSL vaccinated mice conferred the greatest protection in mice that also showed higher induction of IgG2a isotype in the immunized animals. There was notable repression in the bacterial load in the pVSL and S–19 vaccinated mice. Passive transfer of Brucella abortus oligopolysacharides (OPS)–specific monoclonal antibody protected mice from Brucella abortus infection (Arya et al., 1989; Montaraz and Winter, 1986; Philips et al., 1989). Incidentally, it was found that the antibody was of IgG2a phenotype. Similarly, Elzer et al, (2002) has also detected a dominant IgG2a isotype in the infected mice. Our findings also strengthen the observation that antigen specific IgG2a isotype plays important role in eliciting protective immune response against Brucella infection.
CONCLUSION
In conclusion, the present study provides useful information in mounting a strong prophylactic measure against brucellosis by using Cu–Zn SOD and L7/L12 ribosomal protein as combined antigen. Delivery of antigen as DNA vaccine followed by boosting with protein antigen proved beneficial in eliciting protective immunity in murine model.
ACKNOWLEDGEMENT
This work was supported by a research grant (BT/PR3332) from Department of Biotechnology (DBT), New Delhi, India. We are thankful to the Director, IVRI for providing necessary facilities.
REFERENCES
Al–Mariri A, Tibor A, Mertens P, DeBolle X, Michel P, Godefroid J, Walravens K and Letesson JJ (2001). Protection of BALB/c mice against Brucella abortus 544 challenge by vaccination with bacterioferritin or P39 recombinant proteins with CpG oligodeoxynucleotides as adjuvant. Infect. Immun. 69: 4816–4822.
http://dx.doi.org/10.1128/IAI.69.8.4816-4822.2001
PMid:11447155 PMCid:PMC98569
Arya, L. N., Elzer, P. H., Rowe, G.E., Enright, M. F. and Winter, A. J. 1989. Temporal development of protective cell–mediated and humoral immunity in BALB/c mice infected with Brucella abortus. J. Immunol. 143: 3330–37.
Bachrach G, Banai M, Bradstein S, Hoida G, Genizi A and Bercovier H (1994). Brucella ribosomal protein L7/L12 is a major component in antigenicity of brucellin INRA for delayed–type hypersensitivity in Brucella sensitized guneia pigs. Infect Immun 62: 5361.
PMid:7960115 PMCid:PMC303276
Bricker BJ, Tabatabai LB, Judge BA, Deyoe BL and Mayfield JE (1990). Cloning, expression, and occurrence of the brucella Cu–Zn superoxide dismutase. Infect. Immun. 58: 2935–2939.
PMid:2201639 PMCid:PMC313590
Brooks–Worrel BM and Splitter GA (1992). Antigen of Brucella abortus S19 immunodiminant for bovine lymphocyte as identified by one and two–dimensional cellular immunoblotting. Infect. Immun. 60: 2459–2464.Cardena AP, Herrera DM, Zamora JLF, Pina FB, Sanchez BM, Ruiz EJG, Williams JJ, Alvarez FM, Castro RF (2009). Evaluation of vaccination with Brucella abortus S19 vaccine in cattle naturally infected with brucellosis in productive systems found in the Mexican Tropic. Int J Dairy Sci. 4: 142–151.
Cassataro J, Estein SM, Pasquevich KA, Velikovsky CA, Barrera S, Bowden R, Fossati CA and Giambartolomei GH (2005). Vaccination with the recombinant Brucella outer membrane protein 31 or a derived 27 amino acid synthetic peptide elicits a CD4+ T Helper 1 response that protects against Brucella melitensis infection. Infect. Immun 73: 8079–8088.
http://dx.doi.org/10.1128/IAI.73.10.6537-6546.2005
http://dx.doi.org/10.1128/IAI.73.12.8079-8088.2005
PMid:16299302 PMCid:PMC1307072
Cassataro,J, Carlos A. Velikovsky, Laura Bruno, Silvia M. Estein, Silvia de la Barrera, Raúl Bowden, Carlos A. Fossati, Guillermo H. Giambartolomei (2007). mproved Immunogenicity of a Vaccination Regimen Combining a DNA Vaccine Encoding Brucella melitensis Outer Membrane Protein 31 (Omp31) and Recombinant Omp31 Boosting. Clin Vaccine Immunol. 14: 869–874.
Confer AW, Tabatabi LB, Deyoubi BL et al. (1987). Vaccination of cattle with chemically modified and modified and unmodified salt– extractable protein from Brucella abortus Vet. Microbiol. 15: 325–339.
http://dx.doi.org/10.1016/0378-1135(87)90020-4
Corbel MJ (1997). Brucellosis an overview. Emerg. Infect. Dis., 3: 213–221.
http://dx.doi.org/10.3201/eid0302.970219
PMid:9204307 PMCid:PMC2627605
Elzer PH, Smith J, Roffe T, Kreeger T, Edwards J and Davis D (2002). Evaluation of Brucella abortus strain RB51 and strain 19 in pronghorn antelope. Ann N Y Acad Sci, 969: 102–105.
http://dx.doi.org/10.1111/j.1749-6632.2002.tb04359.x
PMid:12381572
Jimenez de Bagues MP, Elzer, PH, Jones SM, Blasco JM, Enright FM, Schurig GG and Winter AJ (1994). Vaccination with Brucella abortus rough mutant RB51 protects BALB/c mice against virulent strains of Brucella abortus,Brucella melitensis,and Brucella ovis. Infect.Immun. 62: 4990–4996
PMid:7927779 PMCid:PMC303217
Ko J and Splitter G (2003). Molecular host pathogen interaction in brucellosis: current understanding and future approaches to vaccine development for mice and human. Clin. Microbiol. Rev.16: 65–78.
http://dx.doi.org/10.1128/CMR.16.1.65-78.2003
PMid:12525425 PMCid:PMC145300
Mallick AI, Singha H, Chaudhuri P, Nadeem A, Khan SA, Dar KA and Owais M (2007). liposomised recombinant ribosomal L7/L12 protein protects BALB/c mice against Brucella abortus 544 infection. Vaccine 25: 3692–3704.
http://dx.doi.org/10.1016/j.vaccine.2007.01.066
http://dx.doi.org/10.1016/j.vaccine.2007.09.008
Montaraz JA and Winter AJ (1986). Comparison of living and non–living vaccines for Brucella abortus in BALB/c mice with O–polysaccharide–specific monoclonal antibodies. Infect. Immun. 51: 961–963.
PMid:3005173 PMCid:PMC260994
Munoz–Montesino C, Adrews E, Rivers R, Gonzalez–Smith A, Moraga–Cid G, Folch H, Cespedes S and Onate AA (2004). Intraspleen delivery of a DNA vaccine coding for superoxide dismutase (SOD) of Brucella abortus induces SOD–specific CD4+ and CD8+ T cells. Infect. Immun. 72: 2081–2087.
http://dx.doi.org/10.1128/IAI.72.4.2081-2087.2004
PMid:15039330 PMCid:PMC375181
Nicoletti P (1990). Vaccination. In: Nielsen, K., Duncan, J.R. (Eds.), Animal Brucellosis. CRC Press, Boca Raton, pp. 284–299.
Oliveira SC and Splitter GA (1994). Subcloning and expression of Brucella abortus L7/L12 ribosomal gene and T lymphocyte recognition of the recombinant protein. Infect Immun 62: 5201–5204.
PMid:7927808 PMCid:PMC303250
Oliveira SC and Splitter GA (1996). Immunization of mice with recombinant L7/L12 ribosomal protein confers protection against Brucella abortus infection. Vaccine 14: 959–962.
http://dx.doi.org/10.1016/0264-410X(96)00018-7
Oliveira SC, Harms JS, Banai M and Splitter GA (1996). Recombinant B. abortus protein that induce proliferation and gamma interferon secretion by CD+4 cells from Brucella vaccinated mice and delayed hypersensitivity in sensitized guinea pigs. Cell. Immunol 172: 262–268.
http://dx.doi.org/10.1006/cimm.1996.0241
PMid:8964089
Oliveira SC, Zhu Y and Splitter GA (1992). Sequences of the rpIJL operon containing the L10 and L7/L12 gene from Brucella abortus. Gene 140:137.
http://dx.doi.org/10.1016/0378-1119(94)90744-7
Oliveria SC, Zhu Y and Splitter, GA (1994). Recombinant L7/L12 protein and gamma irradiated Brucella abortus induce a T helper 1 subset response from murine CD4+ T cells. Immunology, 83: 659–664.
Onate AA, Donoso G, Moraga–Cid G, Folch H, Cespedes S and Andrews E (2005). An RNA vaccine based on recombinant semliki forest virus (SFV) particles expressing the Cu–Zn superoxide dismutase protein of Brucella abortus induces protective immunity in BALB/c mice. Infect. Immun. 73: 3294–3300.
http://dx.doi.org/10.1128/IAI.73.6.3294-3300.2005
PMid:15908354 PMCid:PMC1111843
Onate AA, Vemulapalli R, Andrews E, Schurig GG, Boyle SS and Folch H (2003). A DNA vaccine encoding Cu–Zn superoxide dismutase of Brucella abortus induces protective immunity in BALB/c mice. Infect. Immun. 71: 986–988.
http://dx.doi.org/10.1128/IAI.71.9.4857-4861.2003
PMCid:PMC187304
Philips M, Deyoubi BL and Canning PC (1989). Protection of mice against Brucella abortus infection by inoculation with monoclonal antibodies recognizing Brucella O antigen. Am. J. Vet. Res. 50: 2158–2161.
Schurig GG, RoopII RM, Bagchi T, Boyle S, Buhrman D and Sriranganathan N (1991). Biological properties of RB51; a stable rough srtain of Brucella abortus. Vet. Microbiol. 28: 171–188.
http://dx.doi.org/10.1016/0378-1135(91)90091-S
Singha H, Mallick AI, Jana C, Isore DP, Goswami TK, Srivastava SK, Azevedo VA, Chaudhuri P, Owais M (2008). Escheriosomes entrapped DNA vaccine co–expressing Cu–Zn superoxide dismutase and IL–18 confers protection against Brucella abortus. Microbes Infect. 10: 1089–1096.
http://dx.doi.org/10.1016/j.micinf.2008.05.007
PMid:18602490
Tabatabai LB and Pugh Jr GW (1994). Modulation of immune responses in BALB/c mice vaccinated with Brucella abortus Cu–Zn superoxide dismutase synthetic peptide vaccine. Vaccine 12: 919–924.
http://dx.doi.org/10.1016/0264-410X(94)90035-3
Tatum FM, Detilleux PG, Sacks JM and Halling SM (1992). Construction of Cu–Zn superoxide dismutase deletion mutants of Brucella abortus: Analysis of survival in vitro in epithelial and phagocytic cells and in vivo in mice. Infect. Immun. 60: 2863–2869.
PMid:1612752 PMCid:PMC257246
Unkeless J, Scigliano CE and Freedmen VH (1998). Stracture and function of human and murine receptors for IgG. Annu.Rev.Immunol. 6251–6281.
Vemulapalli R, He Y, Cravero S, Sriranganathan N, Boyle SM and Schurig GG (2000). Overexpression of protective antigen as a novel approach to enhance vaccine efficacy of Brucella abortus strain RB51. Infect. Immun 68: 3286–3289.
http://dx.doi.org/10.1128/IAI.68.6.3286-3289.2000
PMid:10816475 PMCid:PMC97582
Winter AJ and Rowe GE (1988). Comparative immune response to native cell envelope antigen and hot sodium dodesyl sulphate insoluble fraction (P.G) of Brucella abortus in cattle and mice. Vet. Immunology and immunopathology, 18: 149–163.
http://dx.doi.org/10.1016/0165-2427(88)90057-8
Wyckoff JH, 3rd, Howland JL, Scott CM, Smith RA, Confer AW (2005). Recombinant bovine interleukin 2 enhances immunity and protection induced by Brucella abortusvaccines in cattle. Vet Microbiol.111:77–87
http://dx.doi.org/10.1016/j.vetmic.2005.09.004
PMid:16242273
Zhang M, Zhang L, Zhang C, Hong K, Shao Y, Huang Z, Wang S, Lu S (2012). DNA prime–protein boost using subtype consensus Env was effective in eliciting neutralizing antibody responses against subtype BC HIV–1 viruses circulating in China. Hum Vaccin Immunother. 8(11):1630–1637
http://dx.doi.org/10.4161/hv.21648


{kind=link}