
Advances in Animal and Veterinary Sciences


Research Article
Advances in Animal and Veterinary Sciences. 1 (4S): 59 – 64Special Issue–4 (Progress in Research on Viruses and Viral Diseases)
Physicochemical Properties of Bovine Viral Dirrhoea Virus of an Indian Isolate
Satav Jaykumar Satwaji1, Sharda Rakesh1*, Mishra Niranjan2, Rajukumar K2, Tiwari Ashish2, Dubey Shivharan C2
- Department of Microbiology, College of Veterinary Science & Animal Husbandry., Mhow, India
- High Security Animal Diseases Laboratory, Indian Veterinary Research Institute, Bhopal, Madhya Pradesh ,India
*Corresponding author:swaraksha@yahoo.com
ARTICLE CITATION:
Satwaji SJ, Rakesh S, Niranjan M, Rajukumar K, Ashish T, Dubey SC (2013). Physicochemical properties of bovine viral dirrhoea virus of an Indian isolate. Adv. Anim. Vet. Sci. 1 (4S): 59 – 64.
Received: 2013–11–27, Revised: 2013–12–28, Accepted: 2013–12–30
The electronic version of this article is the complete one and can be found online at
(
http://nexusacademicpublishers.com/table_contents_detail/4/190/html
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
Study of physicochemical properties of BVDV revealed that virus can be inactivated at 56oC for 30 min and by the treatment of 1% NaOH, 5% chloroform and 0.05% trypsin for an hour exposure. Ammonium chloride at 10mM concentration completely inhibited the penetration of virus in Sheep Fetal Thymus Riems (SFTR) cell line when treated for 15 min. Methyl blue dye up to 10µM had no effect on BVDV in concentration, however, crystal violet inactivated the virus completely at the same concentration, both in presence and absence of organic matter (milk powder). Acidic pH (≤3) and disulphide bond reducing agent also inactivated BVDV after 1 h exposure. Ethanol was effective at concentrations of 70, 80, and 95 percent in inactivating the virus after 15 sec of exposure at 20oC, whereas propanol inactivated the virus in same time at 30 and 45 percent concentrations. During the past few years, many healthcare workers have changed from antimicrobial liquid soaps to alcohol–based hand rubs for post–contamination treatment of hands as suggested by the CDC guideline for hand hygiene. As BVDV serves as a surrogate model for Hepatitis C virus (HCV), the present study can contribute to evaluate and devise effective methods of controlling the latter.
INTRODUCTION
Bovine Viral Diarrhoea (BVD) is an acute, highly contagious viral disease of ruminants characterized by transient fever, diarrhoea, immunosuppression, fertility problems, thrombocytopenia, mucosal lesions, abortion, neonatal death, congenital malformations, persistent infection of the offspring, podo–dermatitis and recently reported glomerulonephritis (Baker, 1987; Thiel et al., 1996; Goens, 2002; Galav et al., 2007).
The causative agent for BVD, Bovine Viral Diarrhoea Virus (BVDV), belongs to the genus Pestivirus in the family Flaviviridae (Heinz et al., 2000). There are two genotypes of BVDV, BVDV type–1 and BVDV type–2 (Pellerin et al., 1994; Ridpath at al., 1994; Harpin et al., 1995; Van Regenmortel, 2000). In cell culture, pestiviruses appear as two biotypes, known as–cytopathic (cp) and non–cytopathic (ncp) (Kummerer et al., 2000). The ncp biotypes are predominant in nature and are often responsible for majority of the disease syndromes (Nettleton and Entrican, 1995).
Very few studies have been carried out to determine the physico–chemical properties and no studies have been carried out with any Indian isolate. The physicochemical properties and replication characteristics are very important to provide suitable interventions for control of the infection. However, the knowledge regarding the effect of various alcohol–based preparations and disinfectants on BVDV is very limited. Therefore, this study was designed to determine the practical utilities of different decontaminating reagents and methods during laboratory handling of BVDV.
MATERIALS AND METHODS
Place of Work
The study was conducted at the High Security Animal Disease Laboratory (HSADL), IVRI, Bhopal, and Department of Veterinary Microbiology, College of Veterinary Science and Animal Husbandry, Mhow, Madhya Pradesh, India.
Virus Isolate
The BVDV isolate (Ind S–17555), initially isolated from a sheep and available at HSADL, IVRI, Bhopal, was used in this study to investigate the physio–chemical properties of BVDV.
Cell Line
Sheep Fetal Thymus Riems (SFTR) cell line obtained from Riems, Germany and available at HSADL, IVRI, Bhopal, was used in this study. The cells were grown in EMEM medium containing 10% horse serum and were free of any adventitious pestivirus contamination.
Immunoconjugates
Anti–mouse IgG–peroxidase conjugate, raised in rabbits (Sigma, cat.no.A–9044) and anti–bovine IgG (whole molecule) peroxidase conjugate, raised in rabbits (Sigma, cat. No.A–7414) were used in the imunoperoxidase monolayer assay (IPMA).
Effect of Temperature
In this study 1x106 TCID50 dilution of BVDV was used for determining the effect of temperature on its viability. One ml of virus suspension in EMEM was treated at 56oC for 30, 45, and 60 min, respectively in circulating water bath. Then preformed monolayer of SFTR cells in 6 well tissue culture plate was infected with 300 µl of the treated inoculums. The plate was incubated at 37oC for 1h for adsorption of virus. The inoculum was removed, monolayer was rinsed slowly with 500µl EMEM, and 2 ml EMEM supplemented with 2% horse serum (infection medium) was added to the wells. The plate was incubated at 37oC in a CO2 incubator for three to four days.
Immunoperoxidase Momnolayer Assay (IPMA)
After completion of incubation period of three to four days, the plate was tested by IPMA for detection of live/survived virus as per the method described by Wood et al. (2004) with certain modifications. The medium was removed and plate was rinsed with 1X PBS. The cells were then heat fixed by keeping the plate at 80oC for 1h in hot air oven and then allowed to cool to room temperature for 20 min. Cells were rehydrated by adding 300 µl of PBS (1X). Then 50 µl of 1:120 diluted WB112 Mab (specific for NS3 region) was added to each well and incubated at 4oC. After overnight incubation the wells were washed thrice (each of 5 min duration) with 1x PBST (0.05% Tween–20). Then 50µl of HRPO conjugated anti–mouse antibodies (Sigma), diluted to 1:300, was added to each well and incubated for 1 h at 37oC. Again four washings were given with 1X PBST (0.05% Tween–20). Stock chromogen mixture was prepared by dissolving 100mg of AEC (3 amino–9–ethylcarbazole) in 15 ml DMF (N’N’ dimethyl formamide). From this stock, AEC working chromogen substrate was prepared by adding 300 µl of stock chromogen in 5 ml of acetate buffer (pH 4) and 5 µl of hydrogen peroxide (H2O2). 50 µl of this working chromogen substrate was dispensed in each well and plate was incubated for 20 min at room temperature in dark. Rinsing the plate under tap running water stopped the reaction. Development of red color in the cell cytoplasm was considered positive for infectivity / presence of BVD virus, while no color in the cell cytoplasm was considered negative for infectivity or / absence of BVD virus as shown in Figure 1.
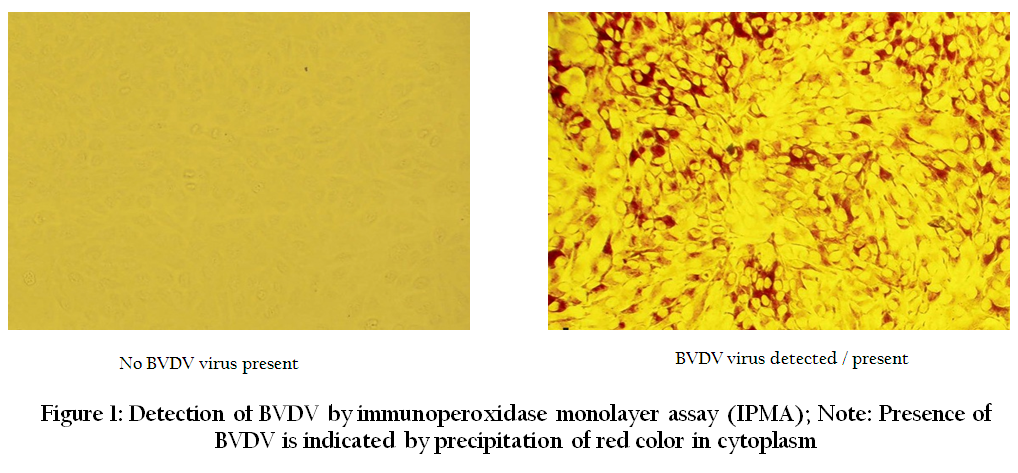
Figure 1: Detection of BVDV by immunoperoxidase monolayer assay (IPMA); Note: Presence of BVDV is indicated by precipitation of red color in cytoplasm
Effect of Dyes
The effect of 1, 5 and 10µM concentrations of methyl blue and crystal violet dyes on BVDV was studied in the presence and absence of organic matter (milk powder). The BVDV was diluted (1104 TCID50 for methyl blue and 1x106 TCID50 for crystal violet) in EMEM. Different concentrations (1, 5 and 10µM) of dye were prepared in 1.5 ml eppendorf tubes by adding 1, 5, 10µl of each of dye (from stock 1mM solution) in corresponding tube and final volume was made up to 1000µl by adding the diluted virus. The material was then kept under artificial light (100 lux) at room temperature for 1h. The preformed monolayer of SFTR cells (3×105 cells/ml) in 6 well tissue culture plate was infected with 300 µl of the treated inoculums and remaining procedure was same as described earlier.
For studying the effect of dyes on BVDV in presence of milk powder, the virus was diluted in EMEM supplemented with 1% milk powder and all other procedures followed were same as described above.
Effect of Alcohols
For studying the effect of alcohols treatment was given as 8 parts alcohol +1 part virus (1x104 TCID50 of BVDV) +1 part medium.
Effect of Ethanol
To study the effect of 70 % ethanol, 80 µl of 87.5% ethanol, 10 µl diluted virus (1x104 TCID50 ), and 10µl EMEM were dispensed in triplicate and mixed properly in an eppendorf tube, while for 80% ethanol 80 µl of 100% ethanol was mixed with identical volumes of diluted virus and EMEM. For 95% ethanol, 90 µl of 100% ethanol and 10 µl diluted virus (1x104 TCID50 ) was dispensed in triplicate and mixed properly. Tubes were incubated at 20oC in a circulating water bath for 15, 30 and 60 sec, respectively and then ice–cold 900 µl EMEM was added in each tube. Subsequently preformed monolayer of SFTR cells in six well cell culture plates was infected using 300 µl of inoculum and remaining procedure was same as described earlier.
Effect of Different pH
One ml of diluted virus (1x106 TCID50 ) in EMEM was taken in each 2 ml eppendorf tubes and then pH of the medium was adjusted to 3, 3.5, 4, 4.5, 5, 5.5, 6, 6.5, and 7 with glacial acetic acid. The material was then kept for 1 h at room temperature and then pH of the inocula was adjusted to 7 by using 0.1M NaOH. Preformed monolayer of SFTR cells in six well plates was infected with 300 µl of treated inoculums and then remaining procedure was followed same as that described earlier.
Effect of DTT at Different pH
DTT is a disulphide bond reducing agent. 450 µl of diluted virus (1x106 TCID50 ) in EMEM and 50 µl DTT (stock 100mM) were dispensed in seven 2ml eppendorf tubes to make final concentration of 10mM DTT in each tube. The pH of the medium was adjusted to 4, 4.5, 5, 5.5, 6, 6.5, and 7 with HCl and kept for 1 h at room temperature. After completion of incubation period the pH of the inocula was adjusted to 7 by using 0.1M NaOH. Preformed monolayer of SFTR cells in six well plates was infected with 300 µl of treated inoculums and then remaining procedure was followed same as described earlier.
Effect of Ammonium Chloride
Different concentrations of ammonium chloride – 1, 2, 3, 4, 5, 6.5, 7.5, 8.5 and 10mM were prepared in 2 ml eppendorf tubes by adding 40, 80, 120, 160, 200µl, 260, 300, 340, and 400µl of ammonium chloride solution, respectively from 50mM stock and final volume was made up to 2000 µl by adding EMEM. The preformed monolayer of SFTR cells in six well tissue culture plates was inoculated with 500µl respective concentration of ammonium chloride and kept at 37oC for 15 min. The media was removed from wells, cells was infected with 300µl of diluted virus (1x104 TCID50 ) and incubated at 37oC for 1 h for adsorption of virus. Then 1.5 ml EMEM containing respective amount of ammonium chloride concentration with 2% horse serum was added in the respective wells. The plate was incubated at 37oC in 5% CO2 incubator for 3 days and rest procedure was followed same as described previously. The virus titration of the cell lysate was carried out by IPMA as per procedure of Wood et al. (2004).
Effect of Sodium Hydroxide
Different concentrations of sodium hydroxide – 0.25, 0.5, 1.0, 1.5, 2.0, 2.5, 3.0, 3.5 and 4.0 percent were prepared by adding 6.25, 12.5, 25, 37.5, 50, 62.5, 75, 87.5 and 100µl of NaOH (from stock 10M NaOH), respectively and final volume was made up to 1000 µl by adding the diluted virus (1x106 TCID50 ). The pH obtained was 8, 9, and 12 for 0.25, 0.5 and 1.0 percent NaOH, respectively, and above 14 for other concentrations. The treated inoculums were kept at room temperature for 1 h and then pH of the inocula was adjusted to 7 by using 0.1 M HCl. Monolayer of SFTR cell in 6 well tissue culture plate was infected with 300 µl of inoculums from the treated material and remaining procedure followed was same as described earlier.
Effect of Trypsin
Different concentrations of trypsin such as, 0.025, 0.05, 0.10, 0.20, 0.25 and 0.3 percent were made by adding 5, 10, 20, 40, 50, and 60µl trypsin (from 5% trypsin stock) respectively, and final volume was made up to 1000 µl by adding diluted BVDV (1x104 TCID50 ). The inuculum was incubated at 37oC for 1 h in a circulating water bath and then horse serum was added to neutralize trypsin. Preformed monolayer of SFTR cells in six well cell culture plate was infected using 300 µl of inoculums from the treated material and remaining procedure followed was same as described earlier.
Effect of Chloroform
In 1.5 ml eppendorf tube different concentrations of chloroform such as 0.25, 0.5, 1, 1.5, 2, 2.5, 3, 3.5, 4, 4.5, 5, 5.5 and 6 percent were made by adding 2.5, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55 and 60µl (from stock of 100% chloroform solution), respectively and final volume was made up to 1000 µl by adding diluted virus (1x106 TCID50 ) in the respective tubes. Preformed monolayer of SFTR cells in six well cell culture plates was infected using 300 µl of inoculum from the treated material and remaining procedure followed was same as described earlier.
RESULTS AND DISCUSSION
Effect of Temperature
The results of the test conducted to assess the effect of temperature (56oC) on virus survivability indicated that BVDV could be inactivated at 56oC for 30 min (Table 1). This has practical implications in that bovine or ovine serum used for cell cultivation can be made free from adventitious BVDV contaminations with such treatment.
Effect of Dyes in Absence and Presence of Milk Powder
After treating the BVDV virus suspension with methyl blue/ crystal violet dye for 1h under artificial light (100lux) at room temperature, the results showed that the methyl blue dye had no effect on inactivating the virus at 1–10 µM concentration either in absence or presence of milk powder (organic matter) as shown in (Table 2). Whereas crystal violet dye has no effect on virus survivability up to the concentration 5 µM either in absence or presence of 1% milk powder. However, crystal violet completely inactivated the virus at and above 10µM concentration both in presence and absence of milk powder as shown in (Table 2). Kevin et al. (2004) studied the effect of methyl blue and methyl violet phenothiazine dyes and reported that both the dyes inactivated the BVDV completely in goat colostrum at 10 and 20µM concentration following 60 min illumination under artificial light. The results correlated well for crystal violet, but not for methyl blue, which may be due to different compositions of the dyes or different virus strains used in the study.
Effect of Alcohols
Ethanol at concentration of 70% and above and propanol at concentration of 30% and above inactivated BVDV completely when exposed for at least 15 sec at 20oC (Table 3).In a study, Gunter et al. (2007) observed that alcohol based hand rubs (ethanol and propanol) reduced the infection of BVD virus by >4log10 steps within 15sec irrespective of the organic load.
Our results corroborated these results and also showed that 70% ethanol and 30% propanol can be assured to be active against clinically relevant enveloped viruses. In the earlier study (Gunter et al., 2007), 75% or more concentration of ethanol has been used while our study included 70% ethanol, which is common hand disinfectant used in many laboratories. We could not evaluate exposure time less than 15 sec but it has only limited clinical relevance.
Effect of Different pH and DTT at Different pH
Treatment of the virus suspension at acidic pH (pH–3 to pH–7) for 1h at room temperature showed that the BVD virus was completely inactivated at acidic pH–3 and below, while it was viable at and above pH–3.5 (Table 4). Whereas the inhibitory effect of DTT (10mM) at various pH levels showed that BVDV was completely inactivated at pH 4.5 and below only (Table 5).
This showed that inactivation of BVDV can take place in presence of acid pH and disulphide bond reducing agent DTT. The survivability at pH 3.5 and above supports earlier evidence of remarkable acid resistance of pestiviruses such as BVDV and CSFV (Krey et al., 2005) unlike alphaviruses and flaviviruses of the family Flaviviridae. This has more practical consequences as BVDV can remain live in meat and meat products of P.I. animals at fairly acidic environment and can be transmitted to susceptible animals. A unique feature of BVDV is the presence of covalently linked homodimers (Erns–Erns, E2–E2) and heterodimers (Erns–E2 and E1–E2), which may contribute to virus resistance to acids (Thiel et al., 1991; Krey et al., 2005).Inactivation of BVDV was only observed when treated with reducing agent DTT at low pH. The inactivation might be due to the reduction of disulfide bonds in the glycoproteins by DTT. The mechanism by which it occurs, however, remains to be elucidated in future studies.
Effect of Ammonium Chloride on Survival and Penetration of BVDV in SFTR cells
As shown in Table 6, it was found that BVD virus was completely inactivated at 10 and 15mM concentration of ammonium chloride, whereas the virus titre was reduced significantly at concentrations of 7.5 and 8.5mM, and there was no effect up to concentration of 3mM.
The effect of chemical inhibitors of endocytosis, such as ammonium chloride on internalization of BVDV has been studied earlier (Krey et al., 2005), which suggest that BVDV invades the host cell via receptor mediated clathrin dependent endocytosis. Ammonium chloride is lysosomotropic weak base, which reversibly neutralize acidification in the endosome and thereby can block completely BVDV infection of susceptible cells. Ammonium chloride at 5mM concentration decreased, upto 6 times, viral penetration into treated MDBK cells. Our study also showed almost similar results, wherein 10 times decrease in viral penetrations was noticed at 5mM ammonium chloride concentration. Whereas BVD virus penetration into treated SFTR cells was completely inhibited at ammonium chloride concentration of 10mM and above (Table 7), which is an important advancement of knowledge regarding adsorption and penetration of BVDV.
Effect of Sodium Hydroxide
As shown in Table 8, after treating the virus suspension at different concentration of NaOH for 1h and after testing resulting cell lysate by IPMA, it was found that NaOH was ineffective at concentration of 0.25 and 0.50 percent (pH–8 & pH–9), while it completely inactivated the virus at a concentration of 1% and above (pH 12) exposed for 1h at room temperature. Also reports of says that NaOH at 0.1M concentrations was shown to inactivate the BVD virus to undetectable level at 60oC in 15s (Borovec et al., 1998). As NaOH solution is widely used for decontaminating biological and laboratory waste, 1% solution can be used for inactivating enveloped viruses such as BVDV, Hepatitis–C and Dengue virus.

Effect of Trypsin and Chloroform
Trypsin inactivated the virus to undetectable level at concentration of 0.05% and above when exposed for 1h at 37oC (Table 9), while, no inactivation was observed with 0.025% trypsin. However, chloroform completely inactivated the BVD virus at concentration of 5% and above after 1h exposure at room temperature (Table 10). This suggests that trypsin and chloroform could be effective virucidal agents against pestiviruses.
CONCLUSIONS
Study of physicochemical properties of BVDV revealed that pestiviruses are inactivated by various physical and chemical agents such as, temperature at 56oC for 30 min, NaOH 1% for 1h, trypsin 0.05% for 1h, chloroform 5% for 1h; Ethanol 70% within 15 sec; Propanol 30% within 15 sec, Crystal violet 10µM for 60 min, ammonium chloride for 15 min exposure, respectively and pH 3 and below after 1h exposure. Propanol (30%) can be used as an effective substitute of ethanol (70%) as a hand disinfectant in laboratory, when latter is not available due to legal restrictions.
REFERENCES
Borovec, S., Broumis, C., Adcock, W., Fang, R. and Uren, E. (1998). Inactivation kinetics of model and relevant blood–borne viruses by treatment with sodium hydroxide and heat. Biologicals, 26: 237–244.
http://dx.doi.org/10.1006/biol.1998.0146
PMid:10208725
Baker, J. C. (1987). Bovine viral diarrhea virus: a review. J. Am. Vet. Med. Assoc., 190:1449–1458.
PMid:3038804
Galav, V., Mishra, N., Dubey, R., Rajukumar, K., Pitale, S. S., Shrivastav, A. B., and Pradhan, H. K. (2007). Pathogenicity of an Indian isolate of bovine viral diarrhea virus 1b in experimentally infected calves. Res. Vet. Sci. 83: 364–368.
http://dx.doi.org/10.1016/j.rvsc.2007.01.019
PMid:17383693
Goens, S.D. (2002). The evolution of bovine viral diarrhea: a review. Canad. Vet. J., 43:946–954.
PMid:12561689 PMCid:PMC339916
Gunter, K., Steinmann, J.and Rabenau, H. (2007). Suitability of vaccinia virus and bovine viral diarrhoea virus (BVDV) for determining activities of three commonly used alcohol based hand rubs against enveloped viruses. BMC Infect. Dis., 7: 5.
http://dx.doi.org/10.1186/1471-2334-7-5
PMid:17291338 PMCid:PMC1803789
Harpin, S., Elahi, S.M., Cornaglia, E., Yolken, R.H. and Elazhary, Y. (1995). The 5' untranslated region sequence of a potential new genotype of bovine viral diarrhea virus. Arch. Virol. 140: 1285–1290.
http://dx.doi.org/10.1007/BF01322754
PMid:7646359
Heinz, F.X., Collett, M.S. and Purcell, R.H. (2000). Classification and Nomenclature of viruses. In: Van Regenmortel MVH, Fauquet CM, Bishop DHL eds. Virus Taxonomy, 7th Rep. Int. Comm. Taxon. Viruses. Publ., Academia Press, California pp 859–878.
Kevin, E., Washburn, R., Streeter, N., Saliki, J. T. and Lehenbauer, T. W. (2004). The use of phenothiazine dyes to inactivate bovine viral diarrhoea virus in goat colostrum. Canad. J. Vet. Res., 68: 105–111.
Krey, T., Thiel, H. J. and Rumenapf, T. (2005). Acid–resistant bovine pestivirus requires activation for pH triggered fusion during entry. J. Virol., 79: 4191–4200.
http://dx.doi.org/10.1128/JVI.79.7.4191-4200.2005
PMid:15767420 PMCid:PMC1061521
Kummerer, B.M., Tautz, N., Becher, P., Thiel, H.J. and Meyers, G. (2000). The genetic basis for cytopathogenicity of pestivirus. Vet. Microbiol., 77:117–128.
http://dx.doi.org/10.1016/S0378-1135(00)00268-6
Nettleton, P.F. and Entrican, G. (1995). Ruminant pestiviruses. British Vet. J., 151:615–642.
http://dx.doi.org/10.1016/S0007-1935(95)80145-6
Pellerin, C., Hurk, J.V.D., Lecomte, J. and Tussen, P. (1994) Identification of a new group of bovine viral diarrhoea virus strains associated with severe outbreaks and high mortalities. Virol., 203:260–268.
http://dx.doi.org/10.1006/viro.1994.1483
Ridpath, J.F., Bolin, S.R. and Dubovi, E.J. (1994). Segregation of bovine viral diarrhea virus into genotypes. Virol., 205:66–74.
http://dx.doi.org/10.1006/viro.1994.1620
PMid:7975238
Thiel, H. J., Plagemann, P.G.W. and Moenning, V. (1996). Pestiviruses. In: B.N. Fields, D. M. Knipe, P.M. Howley (ed.). Fields Virology, 3rd Edn. Lippencott–Raven Publishers, Philadelphia, pp1059–1073.
Thiel, H. J., Stark, R., Weiland, E., Rumenapf, T. and Meyers, G. (1991). Hog cholera virus molecular composition of virions from a pestivirus. J. Virol., 65: 4705–4712.
PMid:1870198 PMCid:PMC248926
Van Regenmortel, M.H.V., Fauquet, C.M. and Bishop, D.H.L. (2000). Virus Taxonomy. Academic Press, San Diego.
Wood, R.D., Goens, S.D., Carman, P.S., Deregt, D., Jefferson, B. and Jacobs, R.M. (2004). Effect on hematopoietic tissue of experimental infection of calves with noncytopathic type 2 bovine viral diarrhea virus. Canad. J. Vet. Res., 68:42–48.
PMid:14979434 PMCid:PMC1142128


